
 Una de las cuestiones a las que se trata de dar respuesta de forma acuciante y concluyente en las disciplinas científicas relacionadas con la biología evolutiva es la de cómo pudo evolucionar la homosexualidad, como comportamiento sexual esporádico o habitual en cualquier sexo de cualquier especie. Dar respuesta a esa cuestión implicaría un estudio amplio y en profundidad de todos los comportamientos susceptibles de ser calificados como homosexuales en toda especie que los manifestase, estudio que necesariamente debería abordarse desde la genética, la embriología, la etología, la neurociencia y todo ello dentro del marco de la Teoría de la Evolución por Selección Natural, que es el paradigma imperante en Biología.
Una de las cuestiones a las que se trata de dar respuesta de forma acuciante y concluyente en las disciplinas científicas relacionadas con la biología evolutiva es la de cómo pudo evolucionar la homosexualidad, como comportamiento sexual esporádico o habitual en cualquier sexo de cualquier especie. Dar respuesta a esa cuestión implicaría un estudio amplio y en profundidad de todos los comportamientos susceptibles de ser calificados como homosexuales en toda especie que los manifestase, estudio que necesariamente debería abordarse desde la genética, la embriología, la etología, la neurociencia y todo ello dentro del marco de la Teoría de la Evolución por Selección Natural, que es el paradigma imperante en Biología.
Cada uno de dichos comportamientos calificables como “homosexuales” (algunos serían de una ambigüedad que dificultaría su clasificación) y en cada una de las especies que lo practicasen, con sus peculiaridades inherentes, podría tener un origen evolutivo distinto. Intentar determinar un origen común a todo comportamiento homosexual de todas las especies sería una empresa arriesgada probablemente condenada al fracaso, puesto que no parecen existir exclusivos determinantes ambientales o exclusivas presiones evolutivas que pudiesen conducir ineludiblemente a su aparición.
Por ejemplo, y hablando de factores ambientales, se conoce que el confinamiento de ciertas especies, sea en cárceles (entre humanos) o en zoológicos (otras especies), pueden hacer aparecer comportamientos sexuales que en estado salvaje raramente se observan. Cuando el entorno poblacional de un animal es predominantemente de su sexo, en algunos casos este puede cambiar su género, cosa que hacen ciertos insectos o peces. No ocurre así con otros muchos animales, cuya sexualidad está claramente definida desde el nacimiento. En éste último caso el impulso a tener relaciones sexuales con miembros de su misma especie y la ausencia de miembros del sexo opuesto podría conducir a la homosexualidad. Pero digamos que estos últimos son casos extraños en la naturaleza, y en cualquier caso bastante forzados.
 Otro ejemplo de homosexualidad, este que respondería en cierto modo a impulsos genéticos, pero no relacionados propiamente con la sexualidad, se daría en especies muy jerárquicas en las que el poder, que generalmente consiste en la precedencia en el acceso a los recursos alimentarios y sexuales reproductivos escasos, se puede expresar ritualmente a través de una cópula homosexual. Entre primates no humanos, por ejemplo, los machos dominantes en ocasiones penetran a los no dominantes, o éstos últimos se les ofrecen como signo de sumisión.
Otro ejemplo de homosexualidad, este que respondería en cierto modo a impulsos genéticos, pero no relacionados propiamente con la sexualidad, se daría en especies muy jerárquicas en las que el poder, que generalmente consiste en la precedencia en el acceso a los recursos alimentarios y sexuales reproductivos escasos, se puede expresar ritualmente a través de una cópula homosexual. Entre primates no humanos, por ejemplo, los machos dominantes en ocasiones penetran a los no dominantes, o éstos últimos se les ofrecen como signo de sumisión.
No hay, empero, ningún factor ambiental o genético conocido que conduzca indefectiblemente a ninguna especie a tener comportamientos esporádicos o habituales de carácter homosexual.
Por ello, en lugar de intentar abarcar demasiado, debemos centrar nuestros esfuerzos en acotar una especie, los comportamientos que en la misma pueden calificarse de homosexuales, el sexo en el que se producen, y la razón que podría encontrarse detrás de la predisposición a este tipo de comportamientos.
El objeto de este post es tratar de encontrar algún posible origen de la homosexualidad masculina en nuestra especie. Acotamos el asunto a la homosexualidad masculina porque estimamos que, aunque puede tener algunos aspectos en común con la femenina es probablemente, a día de hoy, más fácil de intentar explicar que ésta última.
La inquietud que suscita la cuestión está justificada, dado que se trata de “nosotros”, con nuestras complejidades culturales y psicológicas, pero nuestro interés está lejos de tener algo que ver con los aspectos morales o cualquier otro aspecto no científico del problema. Los científicos intentan describir el mecanismo (dentro del paradigma darwinista) a través del cuál pudo la homosexualidad haber evolucionado.
Constituye un aparente escollo a la Teoría de la Evolución por Selección Natural de Charles Darwin que, dentro del linaje evolutivo de una determinada población animal pueda surgir un porcentaje superior al que cabría esperar por azar de individuos orientados sexualmente hacia los individuos de su mismo sexo: esto es debido a que, de darse el caso de una homosexualidad estricta en un conjunto de individuos dentro de una población animal, su orientación sexual y los comportamientos de evitación respecto al otro sexo, darían lugar a una drástica reducción de la descendencia de los individuos de orientación homosexual que, a lo largo de sucesivas generaciones, conduciría a una progresiva eliminación del acervo genético de la población de los genes que predispusieran al comportamiento homosexual.
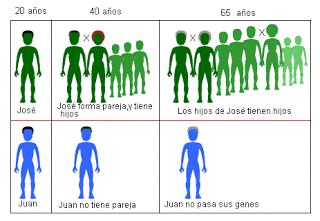 Este planteamiento es probablemente demasiado simple, dado que, en primer término, no todos los homosexuales lo son de forma estricta ni, de hecho, evitan por completo a lo largo de su ciclo vital las relaciones sexuales potencialmente reproductoras con individuos del sexo opuesto. Por otro lado, cualesquiera que fueran los genes que contribuyeran a dar forma a un organismo (y dentro de éste a un cerebro) que generasen conductas homosexuales, no tendrían porqué desaparecer a lo largo de las generaciones, siempre y cuando cada uno de ellos por separado pudiese pasar de una generación a la siguiente a través de portadores no homosexuales o no estrictamente homosexuales. La recombinación genética haría lo demás: los genes que juntos generan una predisposición a la homosexualidad se barajarían con todos los demás con cada nueva generación, dando origen a un porcentaje significativo de individuos con orientación homosexual, y mantenerse en el acervo genético indefinidamente. La homosexualidad, como se ve, no es ni puede ser un callejón sin salida evolutivo, y por tanto su prevalencia en determinados porcentajes en un linaje evolutivo no constituye ninguna paradoja.
Este planteamiento es probablemente demasiado simple, dado que, en primer término, no todos los homosexuales lo son de forma estricta ni, de hecho, evitan por completo a lo largo de su ciclo vital las relaciones sexuales potencialmente reproductoras con individuos del sexo opuesto. Por otro lado, cualesquiera que fueran los genes que contribuyeran a dar forma a un organismo (y dentro de éste a un cerebro) que generasen conductas homosexuales, no tendrían porqué desaparecer a lo largo de las generaciones, siempre y cuando cada uno de ellos por separado pudiese pasar de una generación a la siguiente a través de portadores no homosexuales o no estrictamente homosexuales. La recombinación genética haría lo demás: los genes que juntos generan una predisposición a la homosexualidad se barajarían con todos los demás con cada nueva generación, dando origen a un porcentaje significativo de individuos con orientación homosexual, y mantenerse en el acervo genético indefinidamente. La homosexualidad, como se ve, no es ni puede ser un callejón sin salida evolutivo, y por tanto su prevalencia en determinados porcentajes en un linaje evolutivo no constituye ninguna paradoja.
Dicho esto habría que destacar además la singularidad humana respecto a los comportamientos sexuales en general y los homosexuales en particular. Son muchos los aspectos que nos diferencian en nuestros comportamientos de las otras especies, incluso de nuestros parientes más cercanos, los chimpancés y bonobos. Asimismo las diferencias entre los sexos en sus estrategias cognitivas y conductuales para tener descendencia viable difieren considerablemente, todo lo cual está abundantemente documentado.
Un aspecto de la homosexualidad en nuestra especie que se puede inferir parcialmente de lo anterior (diferencias con otras especies y diferencias entre los sexos), es el de un desarrollo ontogenético del comportamiento sexual que, aun siendo característico de cada sexo particular, resultaría notablemente sensible a señales del ambiente social, con cada generación. Esto tendría su origen en los imperativos que dicho ambiente social ha ejercido filogenéticamente sobre la evolución de nuestra cognición social y de nuestros comportamientos en general, dentro de lo cual los comportamientos reproductivos se pueden considerar una regla antes que una excepción.
Surge, en nuestra especie, la necesidad de un compromiso entre la monogamia y la poligamia, compromiso que adquiere una dimensión nueva (entre las escasas especies potencial o estrictamente monógamas) como consecuencia de nuestras adaptaciones cognitivas al trato social. Sobre el impulso natural al apareamiento se superpone el impulso de proteger nuestra inversión reproductiva, y sobre éste los impulsos de pertenencia al grupo y de búsqueda de crédito y reconocimiento sociales. Ninguno de estos impulsos anula al resto, y en nuestra mente pueden producirse, y de hecho se producen, conflictos a varios niveles que nos llevan a comportarnos, a lo largo de nuestro ciclo vital, con una cierta ambigüedad, cuando no de forma marcadamente contradictoria. La necesidad tiene muchas caras. Y de todo esto puede surgir un precario equilibrio a largo plazo entre monogamia y poligamia, moldeado asimismo por la cultura.
Desde el punto de vista de la orientación sexual particular de cada individuo la cosa se complica un poco, pero no demasiado. El compromiso entre homosexualidad y heterosexualidad es menor, precisamente porque en la evolución, la heterosexualidad es la fórmula más idónea para dejar descendencia. Ello no obsta para que cada individuo, en los genes que afectan de forma directa o indirecta a su conducta sexual, pueda tener una combinación de alelos y una expresión determinada de los mismos que den lugar, con el desarrollo ontogenético, a una conducta marcada por la atracción sexual total o parcial hacia el propio sexo. El contexto social y las vivencias particulares jugarían sobre este desarrollo un papel de extraordinaria importancia. Y la ambigüedad estaría servida en muchos casos. No es fácil determinar el número.
La tesis fundamental que aquí quiero exponer gira en torno a la otra gran aportación de Charles Darwin a una explicación amplia y acerada de la evolución biológica: la Teoría de la Selección Sexual.
En principio podría parecer que ninguno de los miembros de los sexos opuestos podría estar interesado en modificar la orientación sexual de sus potenciales parejas reproductoras. Pero ni la Selección Sexual (más “consciente” que la natural) es algo que opere con claridad diáfana ni sin obstáculos, moldeando su objeto de forma óptima, ni puede ejercerse sobre un particular rasgo fenotípico sin afectar a más de un rasgo genotípico subyacente.
Así, podría haber ocurrido, y de hecho es muy probable que así haya sido, que un sexo seleccionase características fenotípicas múltiples en otro y, colateralmente, otras características asociadas por cercanía genética (física, dentro de los cromosomas o epigenética, o del desarrollo ontogenético, afectando a sus estadios dentro del útero), características estas últimas que no estaban en la mente del “seleccionador” y que, podría decirse, venían en el paquete.
En la evolución de nuestro linaje parecen haber existido algunos hitos particularmente reseñables que hicieron de la crianza de los descendientes un proceso al que había que dedicar cada vez más recursos.
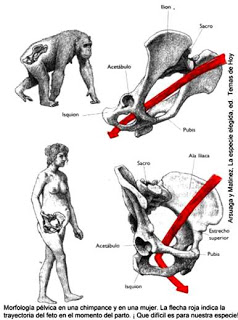 Uno de ellos fue la locomoción bípeda, otro el uso de herramientas, y por último el crecimiento del cerebro. Podemos inferir a partir de los estudios evolutivos actuales que estos hitos se sucedieron en el orden expuesto aquí. Como resultado de unos cambios fueron necesarios, posibles o precisos otros. La liberación de las manos con la locomoción bípeda parece haber sido necesaria para el uso de herramientas, al menos el sofisticado uso que de ellas que nuestro linaje ha venido haciendo. El uso de estas herramientas muy probablemente hizo posible el desarrollo de un cerebro mayor. Y por último, el estrechamiento del canal del parto en las hembras de los homíninos tras los cambios anatómicos generados por la nueva forma de locomoción hizo preciso que una parte importante del desarrollo de un cerebro de mayor tamaño se produjera fuera del útero materno.
Uno de ellos fue la locomoción bípeda, otro el uso de herramientas, y por último el crecimiento del cerebro. Podemos inferir a partir de los estudios evolutivos actuales que estos hitos se sucedieron en el orden expuesto aquí. Como resultado de unos cambios fueron necesarios, posibles o precisos otros. La liberación de las manos con la locomoción bípeda parece haber sido necesaria para el uso de herramientas, al menos el sofisticado uso que de ellas que nuestro linaje ha venido haciendo. El uso de estas herramientas muy probablemente hizo posible el desarrollo de un cerebro mayor. Y por último, el estrechamiento del canal del parto en las hembras de los homíninos tras los cambios anatómicos generados por la nueva forma de locomoción hizo preciso que una parte importante del desarrollo de un cerebro de mayor tamaño se produjera fuera del útero materno.
De esta forma podría explicarse (y así lo hacen muchos estudiosos de la evolución humana, con los debidos matices) que los recursos destinados a la crianza de la descendencia tuvieran que ser cada vez mayores. El esfuerzo de la crianza fue, cada vez en menor medida, una labor exclusiva de las hembras: los machos debían también aportar recursos para la misma, bien directa, bien indirectamente. En la medida en que un macho fuera colaborador las probabilidades de que su descendencia fuera viable eran mayores. Esto, a lo largo de suficientes generaciones podría haber inclinado la balanza de la crianza hacia un mayor equilibrio entre los sexos. Y eso solamente sería consecuencia de la selección natural. Pero, en una cuestión tan central en biología como la de la reproducción y el cuidado de la prole, la selección natural podría haber sido reforzada por las preferencias de unos cerebros femeninos cada vez más sofisticados: ahí entraría en juego, de forma bastante determinante, la selección sexual.
¿Qué rasgos seleccionarían exactamente las hembras de nuestra especie en los machos a la hora de elegir aquel con el cual les resultase más tentador aparearse y tener descendencia? : rasgos de comportamiento asociados a la formación y reforzamiento de vínculos sociales y, especialmente, a la crianza de los hijos. Lo más probable es que las hembras prefiriesen machos con tendencias cognitivas y emocionales hacia las relaciones más estables y cierto gusto por (o mejor sería decir impulso hacia) la crianza de los hijos, tendencias connaturales al sexo femenino pero raramente presente entre los animales machos. Tendencias cuyo sustrato genético y ontogenético estaría en gran medida en el sistema nervioso central.
Por la reducción en la diferencia de tamaño entre los sexos a lo largo de nuestra historia evolutiva cabe deducir que las luchas entre los machos por las hembras fueron cada vez en menor medida la forma de obtención de parejas para el apareamiento. Por otro lado los cerebros de machos y hembras crecieron por igual. Las relaciones sociales, junto con el uso de herramientas, acaso también el nacimiento del lenguaje por la confluencia de ambos fenómenos, convirtieron a los machos de nuestra especie en animales fácilmente “moldeables” por la selección sexual de las hembras. Éstas, que probablemente “domeñarían” o “domesticarían” a los machos en coalición, sólo posible con un cerebro social desarrollado, con sus decisiones tomadas a lo largo de las generaciones, fueron perfilando un “estilo” cognitivo y emocional nuevo en los machos: surgía el compromiso entre monogamia y poligamia de las tensiones evolutivas entre el impulso masculino indiscriminado a dejar descendencia y el impulso femenino a garantizar a la prole una larga y reproductora vida. Se producían asimismo cambios, acaso no muy grandes, en los cerebros y sistemas hormonales masculinos, pero de profundas repercusiones. Para que los machos pensaran y sintieran respecto a sus descendientes de una manera parecida a las hembras, su cerebro y su sistema endocrino tendrían que feminizarse en algún grado….o desmaculinizarse, ya que la opción por defecto del embrión en desarrollo es la femenina. Dicho de un modo que todo el mundo puede comprender, todos somos mujeres con mayores o menores retoques. En el principio era la hembra, y la costilla de Adán no es más que un mito.
Ni la biología ni la evolución funcionan con cambios plenamente ajustados a las necesidades. Así pues, la presión selectiva ejercida por las hembras de nuestra especie sobre los machos para que fueran mejores criadores, afectaron a los genes y al desarrollo masculinos de forma tal que el cerebro masculino pudiera generar comportamientos, cogniciones y emociones parecidos a los femeninos no sólo en lo que respecta al cuidado de la prole, sino también, y de un modo no buscado, a la percepción de la sexualidad.
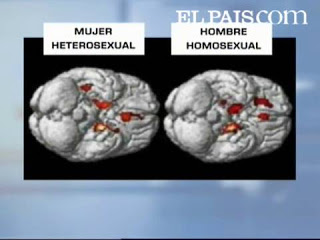 Gracias a la puesta en relación del conocimiento adquirido en embriología sobre cómo la testosterona afecta a los fetos humanos para determinar la sexualidad y ciertos rasgos fenotípicos medibles, como el tamaño del núcleo intersticial 3 del hipotálamo anterior (INAH 3) (LeVay y colaboradores) o la diferencia de longitud entre los dedos índice y anular de nuestras manos, en particular la derecha, se han empezado a identificar algunos de los rasgos que podrían variar entre los sexos en percepción, cognición, emociones, comportamientos…en función de lo expuestos que estuvieron los individuos observados a la testosterona durante un período crítico de su desarrollo embrionario.
Gracias a la puesta en relación del conocimiento adquirido en embriología sobre cómo la testosterona afecta a los fetos humanos para determinar la sexualidad y ciertos rasgos fenotípicos medibles, como el tamaño del núcleo intersticial 3 del hipotálamo anterior (INAH 3) (LeVay y colaboradores) o la diferencia de longitud entre los dedos índice y anular de nuestras manos, en particular la derecha, se han empezado a identificar algunos de los rasgos que podrían variar entre los sexos en percepción, cognición, emociones, comportamientos…en función de lo expuestos que estuvieron los individuos observados a la testosterona durante un período crítico de su desarrollo embrionario.
Por lo que hasta ahora se ha podido comprobar con numerosos sujetos de estudio es que la exposición fetal a la testosterona afecta a numerosos rasgos fenotípicos. Entre ellos uno que parece no guardar relación alguna con el comportamiento sexual o de ningún otro tipo: el Índice Digital D2:D4, esto es, el ratio entre la longitud de los dedos anular e índice. Parece que esta relación puede ser un marcador bastante fiable de la exposición fetal a la testosterona (Zhengui Zheng, Martin Cohn).
Este rasgo “irrelevante” resulta ser un marcador apropiado. Una vez hecha la medición del Índice D2:D4 en todos los sujetos estudiados se procede a identificar y registrar comportamientos, hábitos, costumbres, tendencias, deseos e intenciones (declarados), y, en definitiva, una larga lista de procesos emocionales-cognitivo-conductuales-sociales y contrastar los resultados de todo aquello susceptible de ser medido directa o indirectamente con mayor grado de aproximación con las cifras ofrecidas por el Índice D2:D4. Y se encuentran correlaciones significativas en varios rasgos (que van desde la percepción espacial, la habilidad lingüística, la empatía, la capacidad atlética, etc etc hasta….el cuidado parental) y que generalmente se considera son más preponderantemente masculinos o más preponderantemente femeninos.
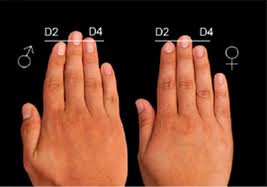 La relación no es perfectamente unívoca e inequívoca. Es decir: los rasgos no van todos juntos de un lado para otro del espectro masculino-femenino (medido a partir del Índice D2:D4), pero sí se correlacionan lo suficiente para pensar que podría, por un lado, existir un sustrato genético que asocie dichos rasgos en alto grado (podría ser una cercanía física en los cromosomas, un interruptor epigenético común….) y por otro que durante el desarrollo embrionario la exposición fetal a la testosterona pueda ir, digamos, por zonas. Probablemente la que más nos interese no sean las manos, aunque como decíamos sirvan de indicador sus dedos índice y anular y la proporción que guardan, sino el cerebro en desarrollo. La testosterona puede inundar el cuerpo, afectando a las gónadas, por ejemplo, o al sistema endocrino, también, pero la importancia que tiene el grado en que afecte al cerebro y las zonas del mismo que queden más expuestas a la hormona es decisiva en la predisposición de la persona aún no nacida pueda tener, a lo largo de su ciclo vital, a sentirse atraída por otras personas de su mismo sexo.
La relación no es perfectamente unívoca e inequívoca. Es decir: los rasgos no van todos juntos de un lado para otro del espectro masculino-femenino (medido a partir del Índice D2:D4), pero sí se correlacionan lo suficiente para pensar que podría, por un lado, existir un sustrato genético que asocie dichos rasgos en alto grado (podría ser una cercanía física en los cromosomas, un interruptor epigenético común….) y por otro que durante el desarrollo embrionario la exposición fetal a la testosterona pueda ir, digamos, por zonas. Probablemente la que más nos interese no sean las manos, aunque como decíamos sirvan de indicador sus dedos índice y anular y la proporción que guardan, sino el cerebro en desarrollo. La testosterona puede inundar el cuerpo, afectando a las gónadas, por ejemplo, o al sistema endocrino, también, pero la importancia que tiene el grado en que afecte al cerebro y las zonas del mismo que queden más expuestas a la hormona es decisiva en la predisposición de la persona aún no nacida pueda tener, a lo largo de su ciclo vital, a sentirse atraída por otras personas de su mismo sexo.
Lo que esto significa es que un hombre puede ser más femenino en alguno de esos rasgos pero poco o nada en otros.
 Las hembras homíninas, con su selección de rasgos más femeninos en los machos de su especie lograron el objetivo de unos padres más proclives a la monogamia y al cuidado de la prole, si bien seleccionaron también a machos cuyos circuitos neuronales relacionados con la atracción sexual tendría un sustrato genéticamente próximo al de ellas, y una exposición ambiental intrauterina durante el desarrollo embrionario que podría comenzar a inclinar la balanza de sus tendencias sexuales hacia las femeninas. Según la cascada de testosterona fuera mayor o menor y según qué zonas del cuerpo en general y el cerebro en particular regase más abundantemente, y por supuesto según que “simiente” (genotipo) se regara, así sería la cosecha. Se mire como se mire la evolución de la homosexualidad masculina como efecto colateral de la selección sexual femenina homínida de machos más fieles y pendientes de su descendencia parece ser una posibilidad en absoluto descartable.
Las hembras homíninas, con su selección de rasgos más femeninos en los machos de su especie lograron el objetivo de unos padres más proclives a la monogamia y al cuidado de la prole, si bien seleccionaron también a machos cuyos circuitos neuronales relacionados con la atracción sexual tendría un sustrato genéticamente próximo al de ellas, y una exposición ambiental intrauterina durante el desarrollo embrionario que podría comenzar a inclinar la balanza de sus tendencias sexuales hacia las femeninas. Según la cascada de testosterona fuera mayor o menor y según qué zonas del cuerpo en general y el cerebro en particular regase más abundantemente, y por supuesto según que “simiente” (genotipo) se regara, así sería la cosecha. Se mire como se mire la evolución de la homosexualidad masculina como efecto colateral de la selección sexual femenina homínida de machos más fieles y pendientes de su descendencia parece ser una posibilidad en absoluto descartable.
La tentativa de hipótesis de toda esta larga perorata sería pues la siguiente:
La selección sexual femenina de hombres con atributos más acordes con la crianza de los hijos y la monogamia (con mayor predisposición a dedicar gran parte de sus recursos a la familia) logró en parte su objetivo al conseguir mayor proporción de machos más «domésticos», pero lo hizo afectando a los genes y a la expresión de los genes relacionados con la sexualidad (dado que los comportamientos de cuidado de la prole son en mucho mayor grado femeninos, estaban asociados por tanto a la sexualidad) entre ellos los que dan forma al cerebro y dentro del mismo a la circutería que determina la atracción sexual. En definitiva: más domesticación conlleva mayor proporción de machos homosexuales.
Harían falta evidencias experimentales. Se trata de una tentativa de hipótesis con demasiadas cosas por contrastar, y como tal la presento. Se pueden derivar de ella algunas predicciones de carácter general:
 1.- Que los homosexuales tienen algunas regiones del cerebro distintas de los heterosexuales, relacionadas con la atracción y el acto de apareamiento.
1.- Que los homosexuales tienen algunas regiones del cerebro distintas de los heterosexuales, relacionadas con la atracción y el acto de apareamiento.
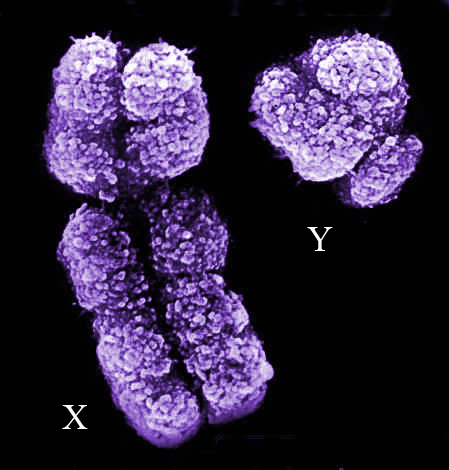
2.-Que los genes que están vinculados a los comportamientos de cuidado parental y elección de pareja exclusiva formarían parte del patrimonio genético femenino e irían asociados con otros (casi todos ellos localizados en el cromosoma X muy cerca unos de otros). La menor expresión del cromosoma sexual X en la pareja XY en los machos podría suponer un comportamiento más agresivo, competitivo, autista etc….más hipermasculino. Pero esta menor expresión podría deberse a los altos niveles de exposición fetal de los embriones masculinos a la testosterona, y por tanto cuanto menores fueran los niveles de testosterona durante el desarrollo embrionario en el período crítico y en lo que se refiere a la parte de este desarrollo que afectara a la conformación del cerebro, mayor sería la probabilidad de que el cromosoma femenino X se expresara, dando lugar a una conformación cerebral (y de otros lugares de la anatomía) algo más femenina. Así que la reducción de estos elevados niveles de testosterona sacaría del armario del hombre a la mujer que lleva dentro, pero según cuánta reducción y en qué momento exacto y según dónde se produjera etc.





Sin duda es muy interesante qué es lo que determina la orientación sexual, pero cualquier orientación, ya que me parece que solo buscar el origen de la homosexualidad es como si se asumiera que la heterosexualidad no se desarrollara por toda una serie de razones biológicas, históricas, sociales, psicológicas, etc…
Creo que ese enfoque dejaría de normalizar la heterosexualidad y asumiríamos de manera natural cualquier otra orientación, porque no solo existe el homosexual.
http://ayudapsicologicacognitivoconductual.com/
En principio me parece una hipótesis razonable, pero veo un problema. ¿Cómo llevamos esta hipótesis a las mujeres homosexuales?
Hola Dani,
Esa hipótesis sólo pretende explorar una posible vía a través de la cuál podrían haber evolucionado mecanismos psicológicos que condujesen a una mayor tendencia hacia comportamientos homosexuales en los machos de nuestra especie. No es aplicable a las hembras.
En ocasiones lo que hay que preguntarse es por qué surge la necesidad de preguntarse. Por qué se elige un tema de estudio. Por qué se elige la hipótesis del nacimiento de un comportamiento. Por qué no plantear el momento del nacimiento del celibato como desviación del comportamiento standar animal. En ciertos temas lo interesante es reflexionar por qué y para qué nos lo preguntamos.
Ad hominem.
Es usted uno de esos que preguntan dando por sentadas las respuestas . Todo un científico. Y yo heterosexual monógamo, para más señas.
Cuando tenga algo que aportar al debate vuelva.
Por cierto, el celibato es también un interesante objeto de estudio, aunque no me consta que nadie nazca célibr.
Es un tema muy controvertido, aunque más por razones ideológicas y morales que por lo que se conoce hasta el momento de él.
Su aportación a este debate en su vertiente científica me parece muy interesante.
Gracias Adrián,
Espero que esta humilde contribución sirva de algo en el desentrañamiento de este enigma.
Comenta que «los científicos intentan describir el mecanismo (dentro del paradigma darwinista) a través del cuál pudo la homosexualidad haber evolucionado». Hombre, a lo mejor existen otras aproximaciones no darwinistas que son igualmente científicas. Es posible incluso que una aproximación antropológica más amplia le ayude a explicar y comprender bastante mejor la homosexualidad que la estrechez darwinista. Inténtelo y a lo mejor se lleva una sorpresa.
Oembuchado, quizás pueda ilustrarnos sobre esas otras aproximaciones. Y le aseguro que no pretendo ser irónico.
Me interesan en particular las aportaciones al debate en cuestión, es decir, sobre el origen de la homosexualidad en nuestras especie. Es por acotar.
Jajaja «una aproximación antropológica más amplia». Quizás quiso decir «antropéntrica más abierta a entender al hombre como algo más que un animal»
Yo, Ralf, imagino que los tiros irán por el gran desarrollo del cerebro que hizo surgir la cultura y con ella un sinfín de posibles combinaciones. Antropología Cultural de Salón. Y si es así me quedo con la «estrechez de miras darwinista» o, como algún otro considera «La idea más grande jamás concebida».